2结果与分析
2.1内生放线菌的分离及转化鬼臼毒素菌株的筛选
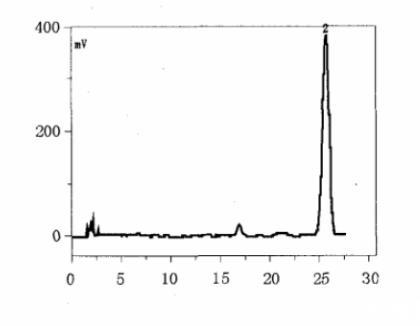
图2鬼臼毒素标准品HPLC图谱
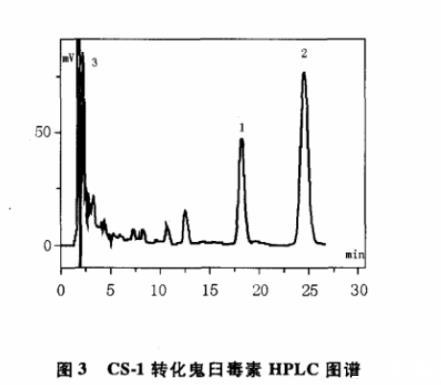
图3 CS-1转化鬼臼毒素HPLC图谱
利用纯培养法从桃儿七根茎分离获得20株内生放线菌,编号为CS-1~CS-20。将20株内生放线菌接种于转化培养基中培养至对数生长期,加入鬼臼毒素继续培养,然后对发酵液进行TCL和HPLC分析,筛选能转化鬼臼毒素的内生放线菌。经过几轮筛选发现,内生放线菌CS-1能转化鬼臼毒素生成产物1。图2是鬼臼毒素标准品的HPLC图谱,图3是CS-1菌株发酵液的HPLC图谱,其中峰1是新出现的峰,为鬼臼毒素转化产物的吸收峰,峰2是鬼臼毒素的吸收峰,峰3是培养基的吸收峰。HPLC检测条件:色谱柱为C18(4.6 mm×250 mm,5μm),流动相乙腈∶水=40∶60,流速1.0 mL/min,紫外检测波长230 nm。
2.2生物转化产物1的结构鉴定
生物转化产物1(15 mg)为白色粉末。ESIMS中m/z 399处出现[M-H]-的分子离子峰,表明产物1的相对分子质量为400,比底物鬼臼毒素小了14,恰好为1个亚甲基。其1H NMR(DMSO)数据与4'-去甲基表鬼臼毒素的文献值一致(表1),可以推断它是4'-去甲基表鬼臼毒。
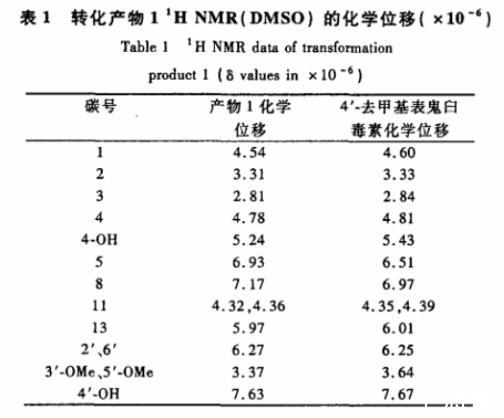
表1转化产物11H NMR(DMSO)的化学位移(×10-6)
2.3鬼臼毒素转化内生放线菌鉴定
2.3.1形态特征菌株CS-1在ISP2培养基中产生基内菌丝和气生菌丝,菌丝无横隔,不断裂(图4)。菌落表面呈皱褶状,颜色呈灰白色;基内菌丝呈黄褐色,不产生可溶性色素。
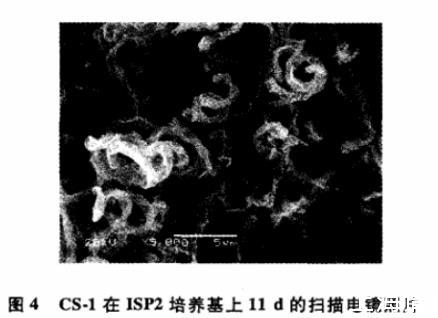
图4 CS-1在ISP2培养基上11 d的扫描电镜照片
2.3.2培养特征菌株CS-1在基础培养基中不长,在蔗糖察氏琼脂培养基中生长较差,ISP3、LB、TSB、ISP4等培养基中生长中等,在其他培养基中生长良好,在这9种培养基中都不产生可溶性色素,菌落形态、基内菌丝颜色变化较大,见表2。
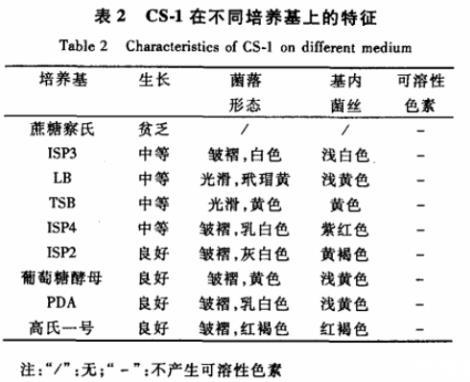
表2 CS-1在不同培养基上的特征
2.3.3生理生化特征
菌株CS-1在20~40℃、pH 6~12条件下可以生长,能耐受10%NaCl,该菌能利用D-阿拉伯糖、D-果糖、D-甘露醇、肌醇、鼠李糖、甘油、胶醛糖、α-半乳糖、D-木糖,不能利用山梨醇、葡糖糖、山梨糖作为碳源;能较好地利用L-缬氨酸、L-谷氨酸、L-酪氨酸,微利用L-组氨酸,不利用L-色氨酸、L-赖氨酸、尿素、甘氨酸。菌株CS-1不能水解淀粉,可使明胶液化,牛奶凝固且冻化,能水解Tween 80、不能分解纤维素;不能使甲基红变红,不产生硫化氢,无法还原硝酸盐,能产生色氨酸酶、氧化酶以及过氧化氢酶。生理生化特征见表3。2.3.416S rRNA的序列分析及构建系统发育N-J树以菌株总DNA为模板,进行目的片段扩增,回收、测序。测序结果(1 434 bp 16S rRNA基因)提交至NCBIGenBank,获得登录号:JQ065718。将菌株16S rRNA基因序列与相近模式菌比对,构建进化树。结果显示,菌株与Streptomyces violaceorubidus LMG 20319T(AJ781374)最为接近,相似度为99.856%;两者在进化树上呈单独1枝,为链霉菌的疑似种,确定为Streptomyce sp.。系统进化树见图5。
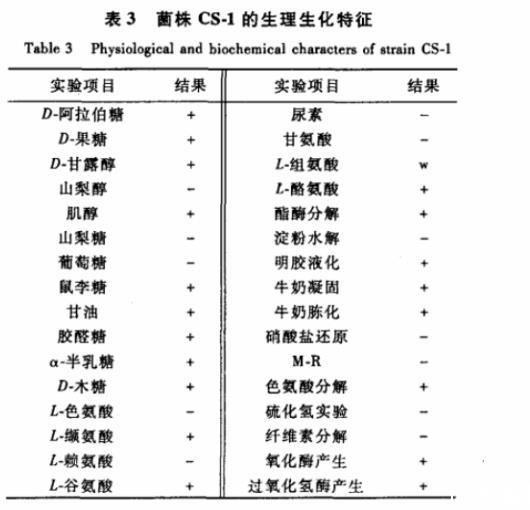
表3菌株CS-1的生理生化特征

图5基于16S rRNA基因序列构建的N-J系统发育树
3讨论
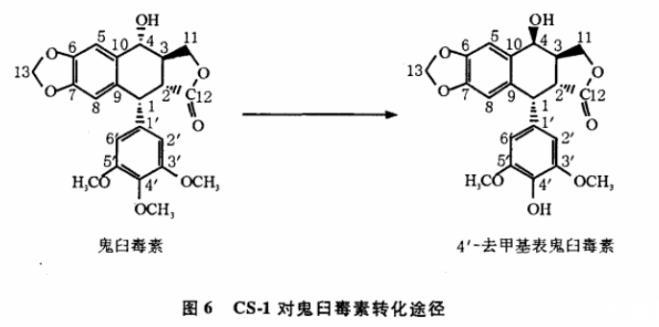
用内生放线菌CS-1对鬼臼毒素进行微生物转化,发现它能将鬼臼毒素转化为4'-去甲基表鬼臼毒素,表明内生放线菌CS-1可能具有O-去甲基化酶,能在鬼臼毒素的甲氧酚上发生去甲基化反应,还可能具有异构化酶将C4位α构型转化为β构型。并且这2种酶是同时或共同转化鬼臼毒素,因为在实验中没有发现4'-去甲基鬼臼毒素或表鬼臼毒素这样的中间体。由此可以推测,内生放线菌CS-1对鬼臼毒素的转化路径可能是由鬼臼毒素直接转化为4'-去甲基表鬼臼毒素(图6)。Hande研究发现4'-去甲基表鬼臼毒素抗肿瘤活性增强,其抗肿瘤机制发生改变,鬼臼毒素是通过破坏正在进行有丝分裂细胞中的微管蛋白集结及微管的形成,使细胞有丝分裂停止在M期,抑制肿瘤细胞正常分裂,发挥抗肿瘤作用,而4'-去甲基表鬼臼毒素是通过抑制DNA拓扑异构酶Ⅱ的活性发挥抗肿瘤作用。因此,可以通过进一步改造4'-去甲基表鬼臼毒素获得新的鬼臼类化合物,为抗肿瘤药物筛选提供更多先导化合物。内生微生物(细菌、放线菌和真菌等)普遍存在于植物组织内,与宿主建立了密切的互惠共生关系。在这种复杂的相互作用中,内生微生物究竟扮演何种角色仍然不清楚。本研究也许能为这种相互作用的分子调控机制的理论探讨,以及转化天然产物的微生物的实验筛选提供一条新的途径。
相关新闻推荐
1、江米酒曲中高产凝乳酶菌筛选、生长曲线及凝乳活力及蛋白水解活力测定(三)