电能细胞是一类能够与外界环境或其他细胞进行电子交换的电活性微生物。其广泛分布于细菌、真菌、古细菌中,目前已分离筛选出上百种具有电活性的微生物,包括向外界环境释放电子的“产电细胞”,以及从外界环境获取电子的“噬电细胞”。以电能细胞为核心构建的多种微生物电催化系统,为解决我国能源和环境等重大需求,提供了极具发展潜力的解决方案,也是国际学术研究前沿的热点和新兴领域。
迄今为止,有两种胞外电子传递机制被广泛认可。即通过外膜c型细胞色素或导电纳米线与电极进行直接接触电子传递,以及通过可溶性电子传递载体(如黄素类、吩嗪类、醌类等)与电极之间进行间接地电子传递。奥奈达希瓦氏菌Shewanella oneidensis MR-1作为一种经典的模式电能细胞,主要通过Mtr途径进行直接电子传递和核黄素进行间接电子传递,目前已有不少研究致力于通过合成生物学策略强化S.oneidensis MR-1的胞外电子传递速率。当前的合成生物学改造策略主要致力于拓宽底物利用谱、提升胞内电子池、过表达细胞色素、增强可溶性电子载体合成、强化生物膜形成以及增强全局调控等,都起到了提高产电的正向效果。
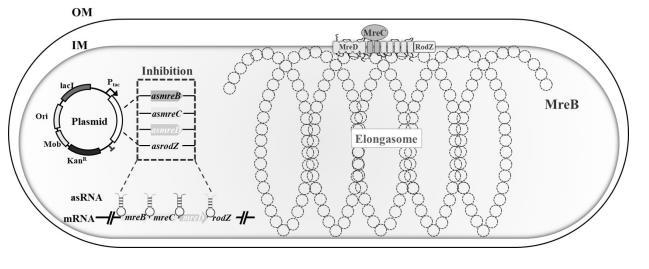
近年来,通过调控基因来改造细菌细胞形态的形态工程成为改进生物制造的一种新方法。这些与细胞形态相关的基因同时参与细胞生长、细胞伸长或细胞分裂,因此细胞形态也受到细胞分裂和细胞伸长等过程的调控。通过形态工程,细菌细胞的形状可以如预期的那样从杆状变化到球形,或者由小到大。这些形态变化随后影响细胞生长代谢速率,从而影响小分子产物合成与分泌、以及包涵体产物的积累和分离。
目前已有很多研究将细菌形态工程用于化学品,如乙偶姻、聚羟基脂肪酸酯和聚乳酸-3-羟基丁酸酯的生产。然而,探究细胞尺寸和细胞能量代谢两间的关系的研究较少。最近的一项研究显示,用细胞分裂抑制剂顺铂处理电活性微生物S.oneidensis MR-1可将杆状细胞转变为细长的丝状形态,这将输出电流密度从野生型111 mA/m2提高到顺铂处理后646 mA/m2。尽管如此,操纵细胞尺寸的方法及其对细胞电生理的具体调控机制尚未建立。
因此,本文运用合成生物学策略对模式产电微生物希瓦氏菌S.oneidensisMR-1进行细胞形态改造,通过表达反义RNA抑制细胞伸长体形成相关基因的表达,调控控制细胞宽度的伸长体,构建了4种不同宽度的工程菌株,通过电化学、生理代谢并结合转录组学从胞内电子产生、胞外电子转移和电活性生物膜形成三个方面,系统地阐明了细胞变宽对电生理的影响。为提高希瓦氏菌的电子传递效率提供有效工程策略。
希瓦氏菌概述
提到发电方式,多数人可能会想到火力、水力、风力、太阳能、核能等途径。然而,微生物发电这一较为冷门的领域却拥有近百年的研究历史。早在1911年,英国生物学家Michael Potter便已经发现,在含有酿酒酵母或特定细菌的培养液中使用铂作为电极,有机物的分解过程可以生成电流。
在生物发电领域,希瓦氏菌(Shewanella oneidensis)MR-1堪称一名“明星”细菌。该细菌最初在Oneida湖中被分离出来,并因其独特的锰还原(Manganese Reducing,简称MR)能力而得名。

希瓦氏菌的表面(来源:《科学》杂志)
在进行锰还原的过程中,希瓦氏菌通过细胞膜上的特殊导电蛋白生成电流,从而获取所需的能量。值得注意的是,该细菌能够在高浓度重金属的厌氧环境中生存,研究还表明它具有将汞离子还原为汞单质,以及将银离子还原为银单质的能力。
那么,希瓦氏菌MR-1为何具有天然的导电特性呢?
科学家将具有自发进化胞外电子转移(Extracellular Electron Transfer,简称EET)途径的微生物称作外产电菌。其中,最受研究者们关注的EET途径便是希瓦氏菌MR-1的金属还原(Metal-reducing,简称Mtr)途径。
在这一途径中,电子通过细胞膜和周质空间内的特殊细胞色素进行传递,这些细胞色素上堆叠着密集的血红素分子。
然而,尽管这些外产电菌在将底物转化为电能的过程中展现出较高的效率,它们对底物(即食物)特别挑剔,适应性相对较差。
此外,由于缺乏有效的遗传操作工具,进行基因改造以提高其应用范围和效率变得相对困难。这些因素在一定程度上限制了包括希瓦氏菌MR-1在内的外产电菌的广泛应用。
为探究反义RNA技术对目的基因的转录抑制水平,工程菌株的转录水平采用实时定量逆转录定量PCR(quantitative reverse transcription-PCR,qRT-PCR)进行验证。
具体操作如下:取工程菌株不同生长时期的二级种子液,在4°C、5000×g离心5min后,用液氮快速冷冻,使用干冰低温寄样交由北京跃丽轩科技有限公司进行qRT-PCR。参比基因选择编码DNA旋转酶的gyrB基因。转录水平通过参比基因gyrB使用2(-ΔΔCT)方法进行归一化处理。
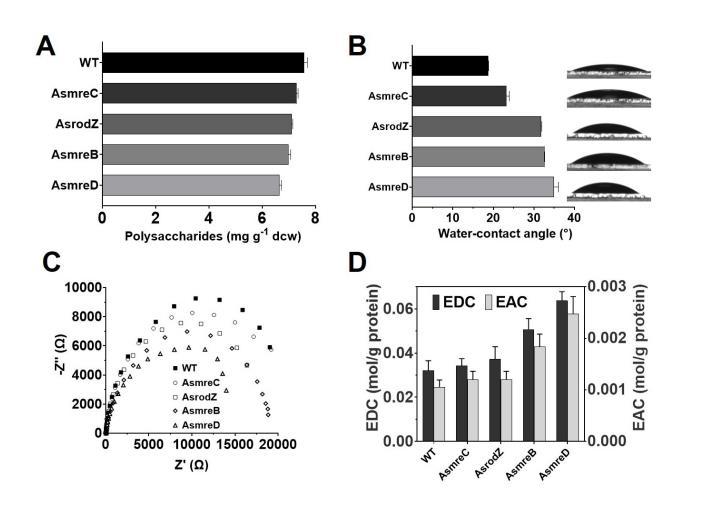
结果显示,工程菌株的具有更高的水接触角,其中菌株AsmreC为23.27°,菌株AsrodZ为31.80°,菌株AsmreB为32.62°,菌株AsmreD为34.98°,均高于WT的18.78°。这是因为工程菌株的细胞表面多糖的减少,导致细胞表面疏水性增强,从而使得水接触角的增大,这有助于生物膜形成初始阶段的表面附着过程。细胞宽度的增加使细胞变得更接近于球状,球状细胞相对杆状细胞更有助于生物膜的垂直扩展,从而增加生物膜的厚度。
电化学阻抗谱扫描表明变宽菌株的生物膜导电性更强,内阻更低。与此同时,变宽工程菌株AsmreB和AsmreD的EDC和EAC明显高于WT,表明工程菌株生物膜的电活性也更高。上述结果表明,胞的扩宽不仅增强了基于c型细胞色素的直接EET,还有助于电活性生物膜形成的初始粘附和垂直扩展过程。
相关新闻推荐
1、新型树枝状多肽非共价递送反义寡核苷酸高效对抗多药耐药细菌感染(上)