球孢白僵菌(Beauveria bassiana)是当前世界上研究和应用最广泛的生防真菌之一,已被开发成为多种制剂用于农林害虫的生物防治。作为典型的丝状昆虫病原真菌,良好的菌丝生长状态是保证球孢白僵菌完整、稳定完成生活史及宿主侵染的重要前提。因此,对影响球孢白僵菌菌丝生长的功能基因研究多年来仍方兴未艾。
生长是球孢白僵菌重要的生防潜能指标之一,影响球孢白僵菌生活史进程,在球孢白僵菌侵染过程中,减缓菌丝生长速率可通过延缓产孢、缓慢侵染从而降低微生物杀虫剂的生防效力。因此,对调控球孢白僵菌生长速率的基因功能研究具有重要的理论及应用意义。近年来,已有众多影响球孢白僵菌生长的基因或蛋白相继被发现,包括MAPK激酶Pbs2、Mkk1和Ste7;热休克转录因子Hsf1、Sfl1以及Skn7;细胞质60S前亚基输出因子BbRei1及WSC1蛋白等,这些蛋白的缺失均引起球孢白僵菌菌落生长速率迟缓。
小GTP酶作为调控细胞信号传导途径的分子开关,行使各种生物学功能,包括细胞极性生长、细胞形态发生、细胞分裂等。在我们之前的工作中,已成功构建球孢白僵菌小GTP酶Ras3和Miro单基因敲除菌株,并检测了它们对球孢白僵菌菌丝生长速率的影响。在SDAY和1/4 SDAY培养基上,ras3缺失菌株Δras3显示出增强的菌丝生长速率,表明Ras3负调控球孢白僵菌菌丝生长速率。与此不同,Miro调控球孢白僵菌细胞内线粒体总量、分布及ATP含量,而Miro功能缺失引起球孢白僵菌菌丝生长速率减缓。在本实验中,成功构建球孢白僵菌Rho5蛋白单基因缺失菌株ΔBbrho5及其对应回补菌株ΔBbrho5∷ΔBbrho5。在SDAY、1/4 SDAY及贫瘠培养基CZA培养基上,ΔBbrho5均表现出受抑制的菌丝生长速率:在富营养培养基SDAY和1/4 SDAY上,ΔBbrho5生长速率相比于野生株下降了10.45%和19.30%;在贫瘠培养基上,ΔBbrho5生长速率相比于野生株下降了9.09%。以上结果表明,小GTP酶对球孢白僵菌生长速率的调节可能是多向的,因基因不同而异,BbRho5正向调控球孢白僵菌菌丝生长速率,这和Miro蛋白功能缺陷引起的表型类似。
提取ΔBbrho5及WT菌株RNA,借助Illumina Hiseq测序平台完成转录组测序并对数据进行分析。
Bbrho5单基因缺失对球孢白僵菌生长速率、多菌灵耐受性及生物防治能力的影响分别刮取野生菌株WT、敲除菌株ΔBbrho5、回补菌株ΔBbrho5∷Bbrho5的分生孢子悬液,分散于吐温-80液体中,将浓度调整为1×107个/mL,悬液均匀涂布于贴有玻璃纸的SDAY平板上,在25℃和光周期12 h∶12 h条件下培养3 d后,用打孔器钻取直径5 mm的菌丝圆片,贴于SDAY、1/4 SDAY或CZA平板上,在25℃和12 h∶12 h条件下培养7 d后,用十字交叉法测量各菌落直径(cm),作为菌落生长速率的指标。
Bbrho5单基因缺失对球孢白僵菌生长速率、多菌灵耐受性及生物防治能力的影响
在富营养培养基SDAY和1/4 SDAY上,ΔBbrho5生长速率相比于野生株下降了10.45%和19.30%。在贫瘠培养基上,ΔBbrho5生长速率相比于野生株下降了9.09%。以上实验结果表明,Bbrho5缺失会引起球孢白僵菌生长速率减缓(图1)。
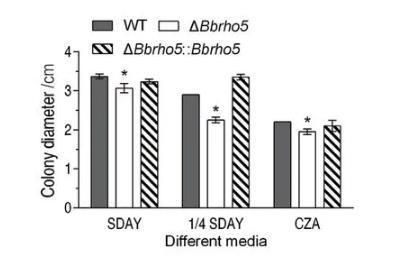
野生菌株、敲除菌株和回补菌株的生长速率测定
此外,Bbrho5缺失对球孢白僵菌多菌灵耐受性及生物防治能力的影响也被检测。结果表明,不同于Bbrho5对生长的显著影响,Bbrho5缺失菌株在多菌灵胁迫条件下,菌落生长直径仅略微下降11%(图2-A)。以大蜡螟幼虫为试虫探究Bbrho5缺失对球孢白僵菌毒力的影响,结果表明,ΔBbrho5对大蜡螟体壁侵染的致死中时LT50相比于野生菌株WT同样表现出略微的延迟(5.07%)(图2-B)。
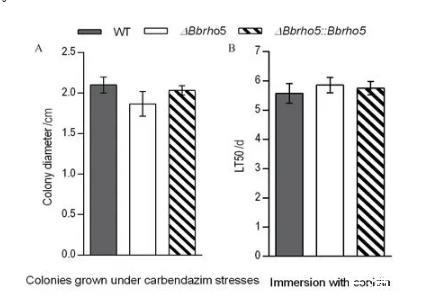
球孢白僵菌对多菌灵的耐药性(A)及对大蜡螟的生物防治能力(B)
结果表明,ΔBbrho5vs WT共获得显著差异表达基因(DEGs)770个,其中显著上调基因有395个,显著下调基因有375个。对ΔBbrho5vs WT基因集中的DEGs进行GO功能富集分析,结果表明DEGs功能主要富集于分子功能部分(molecular function,MF),其中氧化还原酶活力(oxidoreductase activity)和单加氧酶活力(monooxygenase activity)功能极为显著。在生物进程部分(biological process,BP),氧化还原过程(oxidation-reduction process)和跨膜转运(transmembrane transport)最为显著。通过KEGG富集通路分析,结果表明,ΔBbrho5vs WT的DEGs显著差异代谢通路最主要富集在氮代谢(nitrogen metabolism),此外,也有多数基因富集到甘氨酸/丝氨酸和苏氨酸的代谢、丙氨酸/天冬氨酸/谷氨酸代谢、色氨酸代谢、酪蛋白氨基酸、精氨酸生物合成、苯丙氨酸代谢、酪氨酸代谢等氨基酸代谢途径。在微生物细胞中,氮源需要转变为谷氨酸和谷氨酰胺才能进一步被利用,因此,谷氨酸和谷氨酰胺在氮代谢中发挥着重要作用。谷氨酸合酶被证明可正向调控灵芝菌丝生长,谷氨酰胺合成酶是一个重要的氮源调节因子,影响微生物对氮源及能源的利用。在刺糖多孢菌中,谷氨酰胺合成酶基因的敲除同样引发显著的生长抑制。在ΔBbrho5vs WT的DEGs中,富集到氮代谢通路中的功能基因有7个,其中5个上调,2个下调,说明敲除菌株可能采用增强氮源利用以应对Bbrho5缺陷引起的生长迟缓。在上调基因中,包含了重要的谷氨酸合酶(glutamate synthase)基因和谷氨酰胺合成酶(glutamine synthetase)基因,这在转录水平上揭示了ΔBbrho5细胞内谷氨酸合成水平增强,暗示ΔBbrho5可能通过调节上升的氮代谢,进而促进谷氨酸合成,以应对生长速率减弱的生防潜能缺陷。
结论
本研究成功构建球孢白僵菌Bbrho5单基因缺失菌株ΔBbrho5,Bbrho5单缺失能够引起球孢白僵菌的显著生长缺陷,并对球孢白僵菌抗多菌灵能力及其生物防治能力效果极微。转录组数据分析表明ΔBbrho5可能通过氮代谢及多种氨基酸代谢作为响应通路调节菌体的生长速率。