噬菌体是一类感染细菌、真菌、放线菌或螺旋体等微生物的病毒,广泛存在于自然界中。由于噬菌体具有数量多,体积小,繁殖快和基因组的高度可塑性等特点,使得工业发酵过程极易受到噬菌体的感染。噬菌体感染具有普遍性和顽固性,发酵企业一旦受到其感染,损失是不可估量的,并且至今仍无应对良策。
大肠埃希氏菌BL21(DE3)是pET系统的常用表达宿主菌,pET系统使用T7启动子,是有史以来在大肠杆菌中克隆表达重组蛋白的功能最强大的系统,现已作为商品化产品被广泛应用。BL21(DE3)在工厂和实验室经常受到噬菌体感染,因此防治噬菌体感染问题亟需解决。
在细菌和噬菌体的共同进化过程中,细菌在感染过程中通常携带着广泛的防御系统来防止噬菌体裂解。其中,比较典型且目前常用于基因编辑的CRISPR/Cas9系统于2011年在化脓性链球菌中发现,是一种适应性免疫系统,可以保护细菌和古细菌免受噬菌体或质粒等外来遗传元素的影响。目前人为防治噬菌体感染的常见措施主要包括用各种消毒剂杀灭噬菌体控制感染源、菌株轮换、自然突变筛选抗噬菌体菌株等。其中通过的传统的基因工程策略以及基于细菌防御系统的噬菌体防治策略方法由于其操作简单,效果显著而被广泛使用。但是目前各个工厂或者实验室使用的抗噬菌体菌株大多只能抵御某一株或几株特定噬菌体的感染,能够抵御未有噬菌体感染的工程菌株依然值得挖掘。
本文对抗噬菌体菌株大肠埃希氏菌BL21-C和BL21-T进行抗噬菌体能力和生长曲线测定,结果表明通过导入以CRIPSR/Cas9系统为抵御系统的元件pPTCS可赋予BL21-C和BL21-T菌株抵御可以裂解BL21(DE3)的短尾类噬菌体侵染的能力,同时也稳定维持着亲本株的生长情况,说明抵御元件pPTCS的转入并不会影响菌株的生长。
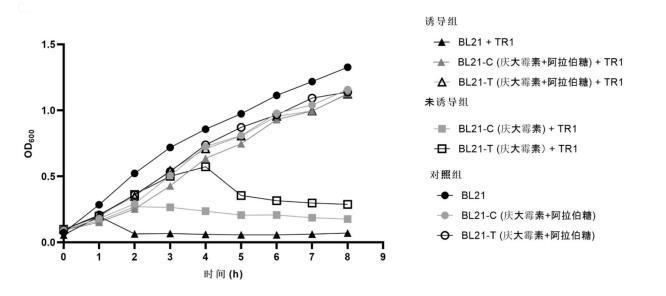
图1为亲本株BL21(DE3)和抗噬菌体菌株BL21-C、BL21-T感染噬菌体TR1(感染复数为1)的生长曲线图。其中,抗噬菌株BL21-C和BL21-T分别测试了在噬菌体TR1感染下有阿拉伯糖诱导和无阿拉伯糖诱导的生长情况。无噬菌体感染设置为对照组。
抗噬菌体菌株BL21-C和BL21-T的生长曲线测定
为测定BL21-C和BL21-T的生长稳定性,设置三组实验:诱导组(噬菌体TR1以感染复数为1侵染且加入终浓度为0.02g/L的阿拉伯糖诱导抵御元件工作)、不诱导组(噬菌体TR1以感染复数为1侵染但无阿拉伯糖诱导)和对照组(无噬菌体侵染)。每组实验设置三组重复。
将BL21(DE3)、BL21-C和BL21-T三株菌株的单克隆分别接种于5mL的LB液体培养基中,其中BL21-C和BL21-T所在培养基中额外加入终浓度为20μg/mL的庆大霉素,诱导组加入终浓度为0.02g/L的阿拉伯糖,在37℃,220rpm的条件下培养至OD600值为0.1后加入噬菌体TR1,该时刻记为0h,随后每隔1h直至8h取样200μL测量OD600值并记录。
结果如图1所示。从图中可以看出,诱导组与对照组的BL21-C和BL21-T生长情况与BL21(DE3)的生长情况基本一致,都呈上升状态。而未诱导组由于未诱导抵御元件工作,没有赋予细菌抵御噬菌体的能力,从而导致其受噬菌体的侵染,OD600值的下降。该结果说明通过转入以CRISPR/Cas9系统为抵御系统的元件,赋予BL21-C和BL21-T菌株抵御噬菌体侵染能力的同时,也稳定维持着原本的生长情况,抵御元件的转入不会影响菌株的生长。
相关新闻推荐
1、福建特产糟制大黄鱼发酵原料红酒糟中短乳杆菌和酿酒酵母生长曲线测定(一)