2结果
2.1重组质粒pJL23-cglⅠ在钝齿棒杆菌中的分离稳定性
重组钝齿棒杆菌在无卡那霉素抗性选择压力的条件下,在LB固体平板上连续传代100次,每隔10代次检测1次质粒丢失情况。实验结果表明,重组质粒pJL23-cglⅠ在重组钝齿棒杆菌中未见明显丢失现象(见表1)。
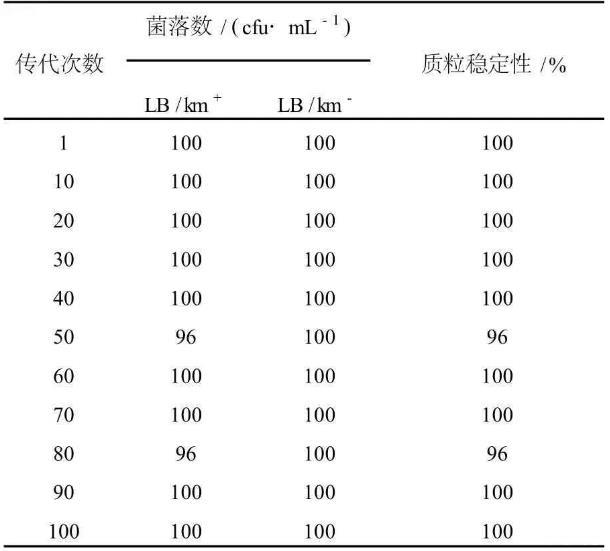
表1重组质粒pJL23-cglⅠ在钝齿棒杆菌中的分离稳定性
2.2重组质粒pJL23-cglⅠ在钝齿棒杆菌中的结构稳定性

表2重组钝齿棒杆菌在不同传代次数中的噬菌体抗性(EOP)
取传代次数分别为原代、20、40、60、80、100次的重组钝齿棒杆菌进行噬菌体抗性定量检测,通过噬菌体感染实验分析cglⅠ基因在传代过程中是否正常表达,以此考察重组质粒pJL23-cglⅠ在钝齿棒杆菌中的结构稳定性。由表2可以看出,重组钝齿棒杆菌在传代过程中一直保持抗噬菌体活性,与原代菌株相比,EOP均小于10-8。表明cglⅠ基因在重组钝齿棒杆菌株中稳定表达,即在传代培养过程中,重组质粒的结构稳定。
2.3重组质粒pJL23-cglⅠ对钝齿棒杆菌生长的影响
从野生钝齿棒杆菌和重组钝齿棒杆菌的生长曲线可以看出(见图1),在相同培养条件下,野生钝齿棒杆菌培养到3 h左右便达到对数生长期,而重组钝齿棒杆菌达到对数期的时间约延迟1 h左右,但二者达到稳定期的时间趋于相同。总体上2个菌株的生长率相差不大,只是重组钝齿棒杆菌进入对数期生长速度要略晚于野生钝齿棒杆菌。
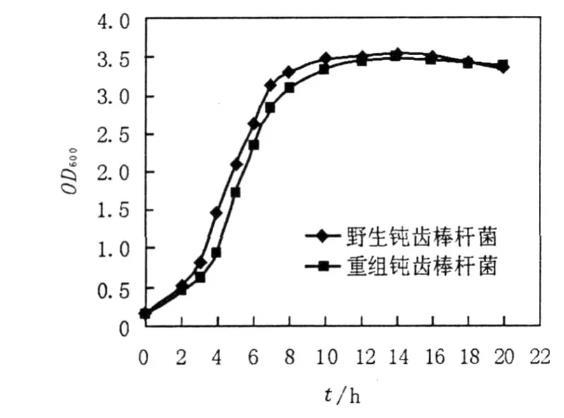
图1野生钝齿棒杆菌和重组钝齿棒杆菌的生长曲线
2.4重组质粒pJL23-cglⅠ对钝齿棒杆菌产生谷氨酸的影响
由图2可以看出,重组钝齿棒杆菌和野生钝齿棒杆菌在发酵过程中谷氨酸积累量的变化趋势基本一致,具有相同的谷氨酸积累高峰,即在发酵32 h有最大的谷氨酸积累量,分别为277.74 mg/L和261.39 mg/L,表明重组质粒pJL23-cglⅠ的导入并未影响钝齿棒杆菌的代谢产酸量。

图2谷氨酸产生动力学曲线
3讨论
对重组钝齿棒杆菌质粒稳定性的研究结果表明,重组菌株连续传100代次后,没有发生质粒丢失现象,且仍具有原代菌株的功能活性,表明重组菌株中的质粒具有结构稳定性和分离稳定性,推测重组质粒稳定性可能与质粒上携带的RM系统cglⅠ基因复合体有关。报道表明,一些Ⅱ型RM基因系统具有分离后杀死活性,可以介导维持质粒稳定性[8-11]。正常情况下,细胞中的M酶保护宿主染色体免遭R酶的致死性裂解。然而,一旦RM基因系统从细胞中被根除,随着细胞分裂,由于稀释,子代细胞中含有的M酶将越来越少,结果是M酶保护新复制的染色体上的许多识别位点的能力渐渐减弱。这样,染色体DNA上未被甲基化的位点将受到攻击,最终导致细胞死亡。同样,RM基因系统丢失后,随着细胞分裂,R酶也将被稀释。然而,M酶和R酶在作用方式上存在不平衡,也就是说,R酶杀死细胞时,只需染色体上一处断裂就可以充分有效(除非DNA被修复),而对于M酶而言,对宿主的保护,则需要染色体上上百个识别位点被甲基化[12]。虽然推测重组菌株的质粒稳定性与质粒上携带的RM基因复合体有关,但其确切原因和机制还有待进一步研究证实。
cglⅠ基因复合体在钝齿棒杆菌中表达后,首先通过甲基转移酶在特异识别序列上使宿主基因组DNA胞嘧啶甲基化,以避免相应限制酶的裂解。这样,宿主基因组DNA胞嘧啶被甲基化后,是否对重组棒杆菌的生长代谢产生影响尚不清楚。本实验结果显示,重组菌株达到对数期时间略晚于野生菌株,表明重组菌株对数期生长有些缓慢,但进入稳定期后两者趋于一致。分析原因可能有以下几方面:①重组质粒在宿主菌中复制、外源基因产物的表达增加了细菌细胞的代谢压力;②含有质粒的重组菌株需要更多的能量和营养合成更多的DNA、RNA和蛋白质等物质;③质粒的复制和表达也将与宿主细胞竞争可用的原料物质,从而影响细菌的生长速率[13-14]。
重组钝齿棒杆菌产生谷氨酸的研究结果表明,重组菌株和野生菌株在实验发酵过程中谷氨酸积累量的变化基本趋于一致,表明重组质粒的导入并未影响菌株的代谢产酸量。
相关新闻推荐
2、牦牛源产气荚膜梭菌qinghai-12的生化特征及生长曲线测定(一)
3、线虫模型:泛藻凝集菌临床与植物有益菌株表型比较研究(二)