对于石生微生物,科学家一直在不断地发现并逐渐深入。由于实验条件的限制,早期的研究一直侧重于其群落结构和微生物多样性方面。最初的研究样品来自于南极的砂岩[4],研究表明微生物定殖形成独特的“地衣结构”[2]。这项研究发表在Science期刊上,对石生微生物的研究有重大意义:①蓝绿藻是石隐生生态系统的初级生产者;②初步阐明了石隐生微生物的定殖方式;③简要探讨了适应极端环境的机制。这三点也是现在绝大多数从事石生生命研究的焦点。
石生微生物的定殖方式
岩石是构成地球岩石圈的主要成分,由一种或多种矿物组成,生物难以分解吸收其中的化学成分。因此,科学家对微生物在岩石的生存方式产生了极大的兴趣:①岩石的内部构造;②石生微生物在岩石内的原位分布。
对于结构十分致密的岩石来说,不存在探讨内部结构的价值。所以科学家将重点关注疏水多孔的砂岩[2,48]和一部分盐岩(石膏)[49-50]。砂岩和石膏都是由单晶体颗粒形成,利用X射线衍射(XRD)可以分析岩石的矿物组成与结构[49]。Wierzchos等[50]利用该技术对石膏的上表面(A)、内部(A′)、底面(A″)的孔隙大小进行了统计分析,发现石膏内部的孔隙明显比外表面的大。石膏内部恰恰是微生物定殖的主要部位,孔隙增大与微生物参与矿物质的溶解有关。

图1 X射线粉末衍射技术对石膏样品的孔径大小分析
利用测量不同位置的光合光量子通量密度(PPFR)的强度,可以对不同深度的蓝藻定殖位置进行假设。再结合环境因子的作用,便可以对石隐生和石下生微生物的原位分布做出模拟。可以看出,整个分布状态可以划分为6层,微生物的定殖部位受到水分、光强、UV辐射、蒸发率以及孔隙度的影响。
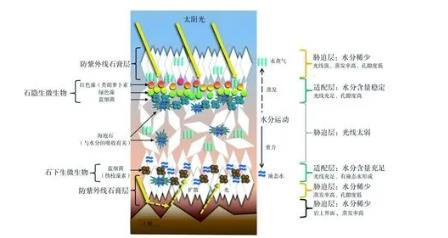
图2阿塔卡玛沙漠石膏样品中石生微生物定殖亚微米级示意图
石隐生微生物在砂岩中形成典型的“地衣结构”,即深(黑)色层为藻类细胞被颜色较深的真菌(地衣共生菌)菌丝包围缠绕形成,白色层为松散的菌丝,绿色层主要为藻类细胞。

图3南极干谷石隐生微生物定殖示意图
石生微生物极端环境适应机制
石生微生物生存的环境条件多种多样,例如干旱、寒冷、高温、盐碱化、UV辐射、寡营养等。在长期的进化过程中,石生微生物形成了耐受各种极端条件的机制。这些机制除了微生物自我调节外,最重要的是岩石本身对微生物适应的帮助是巨大的。
水分:干燥对于生存在荒漠中的石生微生物来说,水是异常珍贵的。大量的蒸腾和稀缺的降水导致生命体可利用的水资源十分稀少。对于石生微生物而言,有着独特的水分吸收方式和耐旱装置。在南极荒漠,雪的融化为生物提供唯一的水源[2,52-53]。在夏天,有少量的雪可以融化从而流入岩石中[54]。岩石基质和地衣结构在高湿度条件下可能会收集和浓缩水[55],紧密的地衣结构和岩石间隙通过物理机制(毛细作用)将融化的雪水或高湿度的水蒸气吸收进石头,此过程并非生物过程。疏松多孔的岩石基质就像水库一样,可以将融水储存起来[2],长达几天甚至数周。南极之外的荒漠,由于早晚温度发生急剧变化,使得一部分水分子凝结附着在石头表面形成露水[30,56],一部分水分子形成雾[57],对微生物而言,都是不可或缺的水资源。有研究表明,盐的潮解也是一种水的潜在来源[58]。例如,当空气相对湿度(RH)>75%时,水蒸气会在NaCl晶体周围凝结成水。当RH<75%(NaCl)的时候,盐岩也可以自发地在周围100 nm的空间内利用岩石颗粒形成的纳米多孔结构(nanoporous structure)的毛细凝聚作用获得液态水[59]。Wierzchos等[42]在室内做石生微生物的湿度控制实验,发现在RH>60%的时候,石膏(内生蓝细菌)会发生水分子自吸现象。所以,盐岩在获取液态水方面有着天然的结构优势,从而确保了石生微生物的正常生长。石生微生物的耐旱形式多种多样。有的细菌可以分裂产生一种特殊的球形细胞,度过多个周期的干湿循环或者长时间的干燥情况[60]。一些黑色的真菌[61]、南极绿藻(卵囊藻科)[62]和所有的蓝细菌[63]具有粗多糖鞘,可以用来保护细胞免受干燥伤害[64]。细菌的孢子(厚壁菌门的内孢子、放线菌的外生孢子)和厚壁孢子(蓝藻)是被记载最广泛地解释细菌脱水耐性的机制,还有报道固氮菌的囊肿和粘球菌的粘孢子也对干旱起抵抗作用[60]。另外,微生物可以合成一些调渗物质(osmoprotectants)来维持细胞活性。例如甜菜碱、蔗糖、海藻糖和四氢嘧啶等。Hershkovitz等[65]从内盖夫沙漠石生样品里分离出1株拟色球藻,可以在渗透胁迫下积累海藻糖和蔗糖。Garcia-Pichel等[66]研究表明,丝状藻青菌在土壤湿润的条件下可以迁移到土壤表面,干燥的时候又会撤退到石头里,这也是微生物应对干旱自我调节的表现。
温度:低温早期研究发现,南极洲石生微生物中真核藻类的生长温度上限在25℃左右[67]。后来发现从南极洲石内分离出来的蓝藻并不是嗜冷菌[68],虽然它们能在4~9℃的环境下生存,但是最适生长温度为20~30℃。Quesada等[69]推测菌群缺乏对极端低温的适应也许是一种优化调节,不仅仅是因为短短几个小时内温度会发生急剧变化,而且极端低温的环境对它们的生理压力也是巨大的,换来的只是高的死亡率。温度最高的夏天,微生物生长代谢也最活跃。-10~5℃之间的岩石温度并不适合微生物生长,但是从群落角度来看,岩石微生物光合固定产量最大,群落达到了最大的适应性[70]。也就是说,相比于单体物种来说,微生物群落拥有适应广泛温度变化的热制度(状况)[71]。最适温度以外的物种,通过调节初级生产者和消费者的比例(Rp/C)达到适应环境的目的。环境温度上升,群落的Rp/C值会升高。Friedmann等[71]在世界范围内的地衣群落内验证了这种猜想。在荒漠环境,细菌对干燥的耐受比对温度的适应更重要[60]。因为只有当岩石温度很低(20℃以下),限制细菌活动的水才有可能出现。石生微生物的耐低温形式也是多种多样。Finegold等[72]从南极洲一个石隐生微生物群落分析得到,细胞膜含有大量多不饱和脂肪酸。纯化得到的磷脂在-20℃依然保持液态,并具有不同寻常的水化功能。电子显微镜显示,膜脂质在温度低于-20℃时,没有形成典型的多层结构,而是形成液体晶体或无序流体状态,维护南极微生物的生存,甚至到-50℃。胞外多糖被认为是细胞外限制水分丧失和细胞内冰晶形成的有效物质[69,73]。一些蓝藻也会合成一些调渗物质,防止干燥和冷冻带来的渗透压伤害[74],如甜菜碱、甘油、海藻糖、甘露醇等相容性溶质[75-76]。一般情况下,冷环境中一个长期性的休眠也是微生物生存能力的体现[69]。南极地衣有其独特的适应方式。它通过改变生长模式,从密丝组织到单丝体,同时保留着恢复叶状体(密丝组织)的潜力,如果条件满足,随时可以恢复到密丝组织结构[2]。
光照:UV辐射、光抑制、光氧化光量子对于石生微生物是有害的。暴露在阳光下的岩石会形成光抑制和细胞损伤、基因组损伤、光合色素漂白、光化学性能降低等[77-78]。除了岩石表层可以降低一些辐射外,石生微生物能合成“反射”紫外线的“屏幕”[64,79-80],例如真菌中的黑色素[61]、蓝细菌或藻类中的类胡萝卜素[81-83]和蓝细菌中的伪枝藻素[81-82,84-85]。其中,类胡萝卜素是吸收紫外辐射而提高细菌耐受性最多的一种色素。在高强度的辐射中生存下来的微生物的耐辐射能力会大大增强,可以达到13 kJ/m2[80]。多细胞聚集体外面形成一层“保护层”,防止过多的辐射渗透到内层细胞。微生物在岩石中定殖的位置和生物量与光合光子通量(PPF)及光合有效辐射(PAR)息息相关。研究表明,只有1%入射光可以穿透砂岩表层,在1.2~3.0 mm深度之间,每毫米就有70%~95%的损失[9]。而且,不同的石质对光合有效辐射的影响很大[30]。例如46.8%的透明燧石中藻类可以生长到40 mm深,厚度范围为5~15 mm,而20.9%的不透明燧石只有30 mm深,厚度范围为5~10 mm。随着入射深度的增加,入射光谱逐渐向红光转变[53],这说明红光在燧石中的穿透力强于短波长的光,所以石生微生物即使在光线暗的情况下也能较好地吸收光能[30]。辐射对石生微生物大分子物质造成很大的光氧化损伤,包括DNA和蛋白质链的断裂等。Daly等[86]发现恐球菌纲和其他的耐辐射菌通过增加细胞内的锰离子浓度,进而降低活性氧浓度来对抗辐射或干燥带来的蛋白质氧化损伤。而且,恐球菌纲中的一些特殊蛋白与RecA、RecFOR等蛋白相互作用对DNA的修复有很大帮助[87-88]。恐球菌纲还有一个高度浓缩的拟核,通过限制游离DNA末端的扩散可以高效地进行DNA修复[89-90]。石生微生物中的共球藻属,还可以调节类胡萝卜素与叶绿素比例增加,这也是一个适应防止光氧化损伤的过程。