2结果与分析
2.1 COR处理番茄叶片后基因表达分析
2.1.1乙烯信号途径相关基因的相对表达量
从图1中可以看出,所有处理组中的EIN2基因和EIN3基因的表达量均显著高于对照组。对于EIN2基因来说,各处理组中表达量最高的为1μmol/L组,相对表达量最低的为0.001μmol/L组,1μmol/L组的表达量显著高于0.1μmol/L组,0.01μmol/L组,和0.001μmol/L组。0.1μmol/L组,0.01μmol/L组和0.001μmol/L浓度的COR诱导EIN2没有显著差异。与EIN2基因表达类似,EIN3基因相对表达量最高的处理组为1μmol/L组,显著高于其他3个处理组,表明COR在高浓度时对EIN2基因和EIN3基因具有较强的诱导作用。
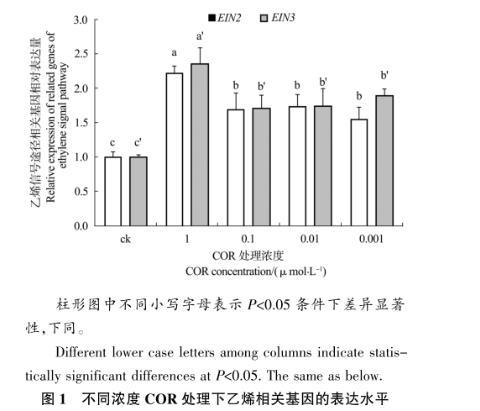
图1不同浓度COR处理下乙烯相关基因的表达水平
2.1.2茉莉酸合成和信号途径相关基因的相对表达量
从图2可以看出,除0.1μmol/L外,其他处理组中的LoxD基因和MYC2基因的表达量均与对照组差异显著,且同一处理组中两个基因的变化趋势相同。1μmol/L COR处理组中的LoxD基因和MYC2基因的表达量显著高于对照组,表明高浓度COR能够诱导番茄叶片表达LoxD基因和MYC2基因。0.01μmol/L COR处理组和0.001μmol/L COR处理组中的LoxD基因和MYC2基因的表达量显著低于对照组,表明低浓度和较低浓度COR处理抑制了番茄叶片LoxD基因和MYC2基因的表达。
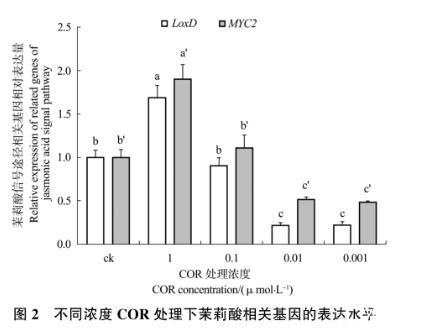
图2不同浓度COR处理下茉莉酸相关基因的表达水平
2.1.3 PR1a基因的相对表达量
从图3中可以看出,除1μmol/L处理组外,其余各处理组PR1a基因的表达量均低于对照组,其他处理组与对照组间差异显著,0.1μmol/L处理组分别与0.01μmol/L和0.001μmol/L处理组均无明显差异,而0.01μmol/L和0.001μmol/L处理组差异显著,整体来说COR能够抑制叶片PR1a基因的表达。0.01μmol/L处理组的PR1a基因表达量在4个处理组中最低,仅为1μmol/L处理组的0.24倍,较低浓度COR对PR1a基因表达的抑制作用最强。
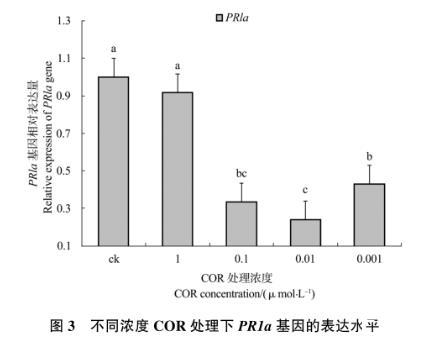
图3不同浓度COR处理下PR1a基因的表达水平
2.1.4 PAL5基因的相对表达量
从图4中可以看出,各浓度COR处理番茄叶片均能诱导PAL5基因的表达。1、0.1、0.01、0.001μmol/L处理组的PAL5基因平均相对表达量分别为307.11、241.76、169.35和171.39。0.1μmol/L处理组分别与1μmol/L和0.01μmol/L处理组之间无明显差异,0.01μmol/L处理组和0.001μmol/L处理组无明显差异。总的来说,随着COR浓度的增高,PAL5基因的表达量呈上升趋势。
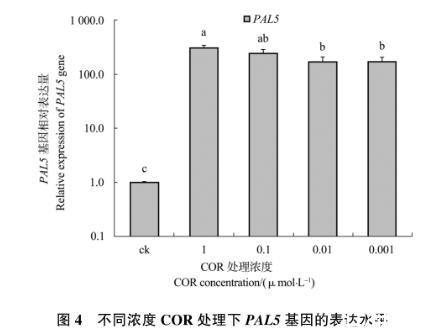
图4不同浓度COR处理下PAL5基因的表达水平
2.2细菌生长能力测定
从图5中可以看出,野生型菌株Pto在番茄叶片中的生长能力极显著高于COR缺失突变体cor-。同一接种浓度下接种不同菌株72 h后,叶片中Pto的种群密度为7.32×107CFU/cm2,cor-的种群密度为1.44×107CFU/cm2,Pto的种群密度约为cor-的5倍,表明病原菌不能分泌冠菌素后,病原菌的生长量明显减少,说明COR能够促进病原菌在宿主植物中的生长。
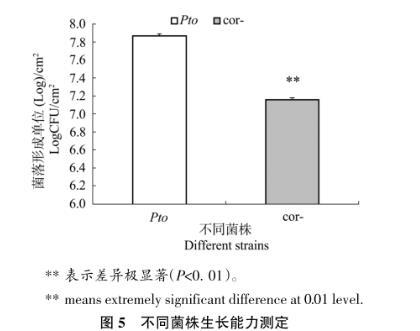
图5不同菌株生长能力测定
2.3胼胝质沉积分析
胼胝质是植物抵御病原菌入侵的物理屏障。从图6中可以看出,处理组诱导的胼胝质沉积数量极显著高于对照组,同时Pto处理组的胼胝质沉积数量极显著低于cor-处理组。荧光染色叶片观测结果表明,接种细菌之后,番茄叶片启动诱导胼胝质沉积的基础防御,而COR促进细菌一定程度上抑制胼胝质沉积,从而抑制植物防御。
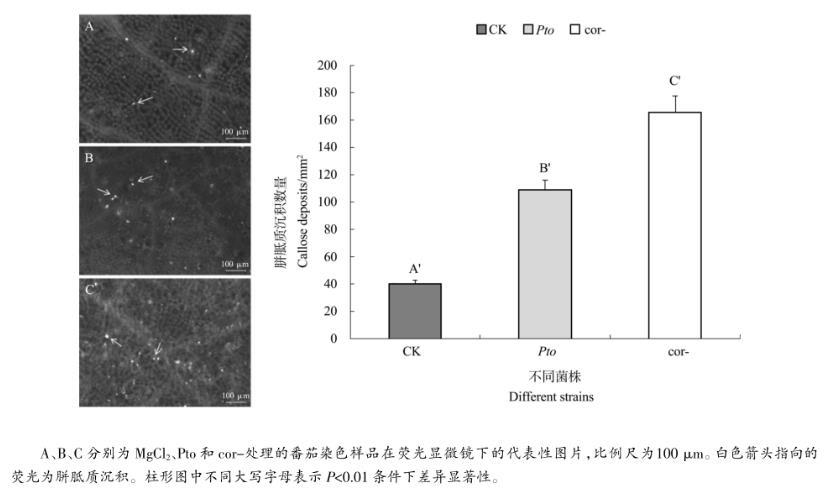
图6不同处理诱导的胼胝质沉积