主要结果
1、通过比较脓毒症患者和HCs之间的微生物群来区分脓毒症相关细菌
首先,我们比较了脓毒症组和HC组之间的α多样性和β多样性。脓毒症患者的肠道α多样性明显低于HCs患者(Shannon指数2.146 vs.2.911,p<0.01;图2A)和不同的微生物结构(ANOSIM检验,p=0.001;图2B)。脓毒症和HCs组流行菌分布如图2C所示。脓毒症患者的变形菌门和肠球菌科数量增加,厚壁菌门和毛螺杆菌科数量明显减少。LefSe发现,脓毒症患者的健康促进菌群(如Blautia、Anaerostipes、Bifdobacterium和Eubacterium_hallii)丰度也显著减少。相反,医院内感染常见的肠球菌(Enterococcus)和克雷伯菌(Klebsiella)在脓毒症患者中的富集程度高于HCs(图2C,D)。此外,与HC相比,脓毒症患者的细菌关系网络不那么复杂,其中肠球菌对其他分类群表现出异常的抑制作用(图2E)。
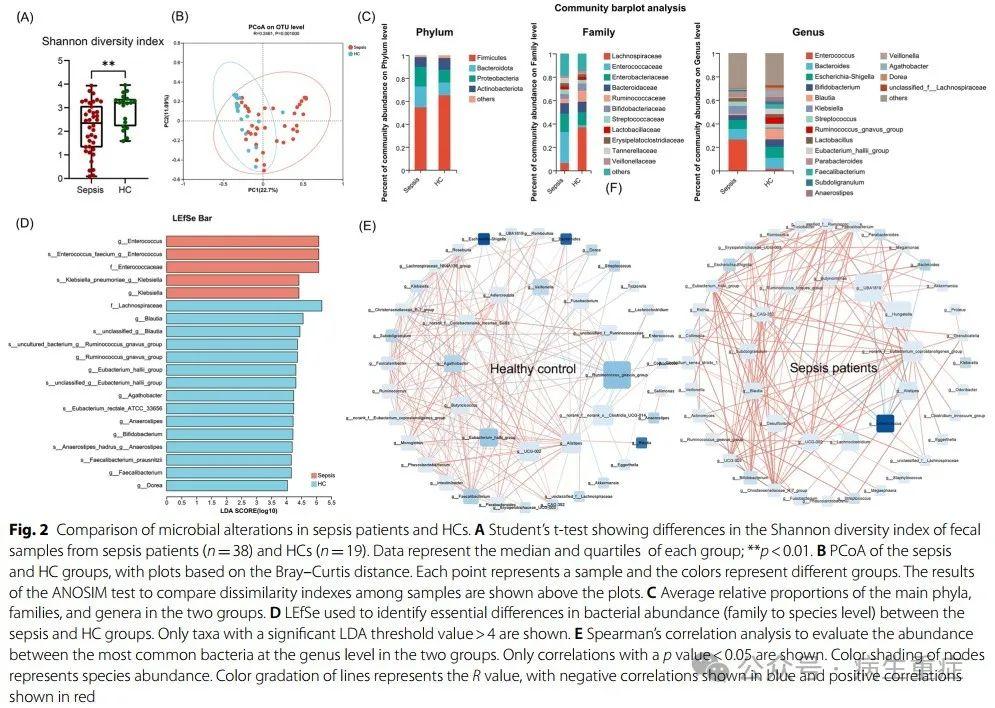
2、脓毒症患者代谢特征与微生物群改变的相关性
接下来,我们对脓毒症和HCs患者的粪便样本进行了非靶向代谢组学分析。共鉴定出449种注释的差异代谢物,脓毒症组中有10种(2.23%)比HC组丰富,表明这些患者的代谢活性可能减弱(图1B和3A)。PCA分析显示,两组患者在正离子和负离子模式下的代谢谱不同(图3B)。我们评估了各种变量对代谢组的影响,发现抗生素的使用数量可能会影响代谢组成。利用基于KEGG注释的代谢富集途径分析,将脓毒症改变的代谢物映射到它们的生化途径(图3C)。在81个潜在的功能差异代谢物中,验证组检测到38个,我们将其定义为脓毒症相关代谢物(图1B)。我们还评估了脓毒症患者中脓毒症相关代谢物与其他参数之间的关系,并认为只有在验证组中证实的相关性是可靠的(图1C)。
差异代谢物似乎与氨基酸代谢密切相关,包括色氨酸、酪氨酸、精氨酸和脯氨酸代谢。参与不平衡的嘌呤代谢、植物次生代谢物的生物合成和ABC转运蛋白也被观察到。考虑到肠道微生物和代谢物之间的密切联系,我们评估了改变的分类群与脓毒症相关代谢物的相关性(图3D)。发现多种细菌与这些关键代谢物密切相关。在前五名中,有四名属于毛螺菌科,表明该细菌家族具有强大的代谢调节功能。值得注意的是,OTU808(Enterococcus faecium)与代谢物几乎呈负相关,这与上述肠球菌与脓毒症肠道微环境中其他细菌的抑制关系类似。此外,我们还鉴定了10种与细菌最密切相关的代谢物:3-甲基二氧吲哚、5-羟基吲哚乙酸、多巴醌、N,N-二羟基苯基丙氨酸、异烟酸、脱氧胆酸、2-羟基肉桂酸、4-羟基苯甲醛、胸腺嘧啶和十六烯二酸。其中6种是芳香化合物,可能来源于芳香氨基酸(AAA)的代谢物,包括色氨酸、酪氨酸和苯丙氨酸。其中,参与色氨酸代谢的代谢物3-甲基二氧吲哚与脓毒症患者肠道微生物的相关性最大。
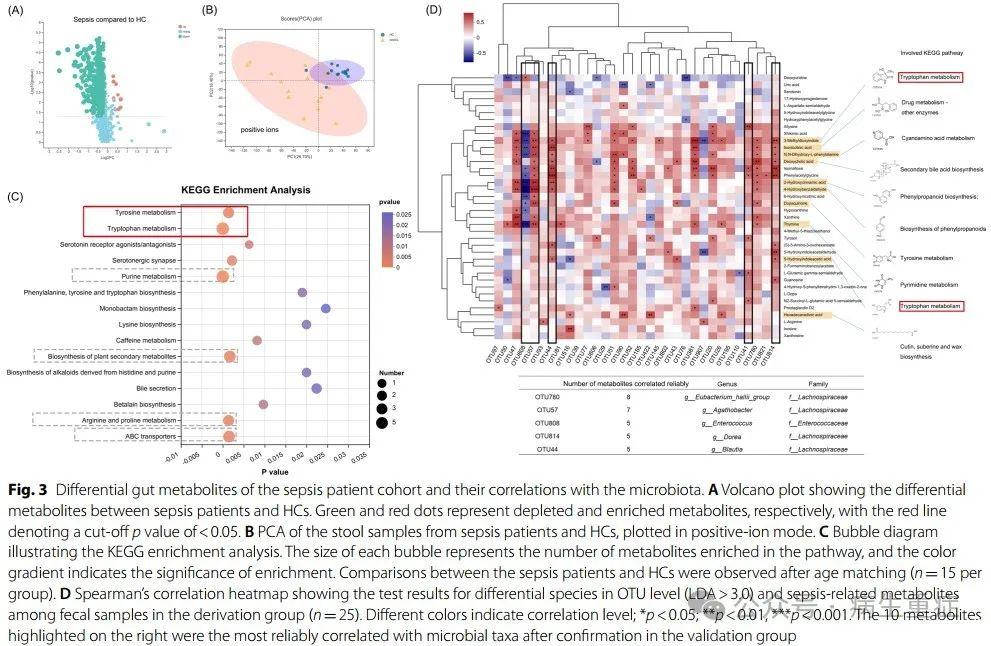
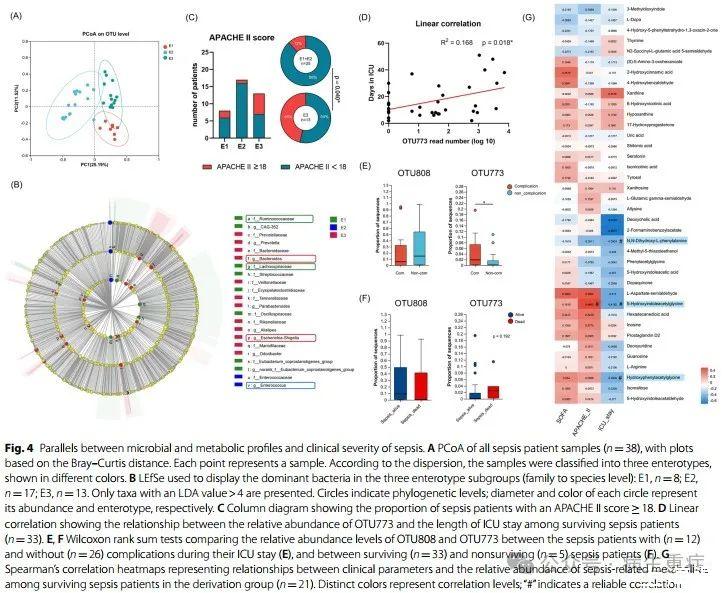
3、肠道微生物和代谢结构与脓毒症临床严重程度的一致性
接下来研究了肠道微生物组和代谢物对脓毒症严重程度的潜在影响。如图4A所示,该组患者根据其肠道微生物组成(也称为肠型)大致可分为三个亚组。在肠型1(E1)中占优势的是毛螺科和瘤胃球菌科;肠型2(E2)以肠球菌为主;肠型3(E3)富含拟杆菌和志贺氏杆菌(图4B)。肠型比较显示,E3期患者最为关键,APACHE II评分≥18的比例明显高于非E3期患者(46%vs.12%,p=0.040;图4C)。此外,E3组患者SOFA评分≥10分、ICU住院≥30天的比例可能更高,死亡率也最高,但差异无统计学意义。尽管E2组患者细菌多样性和丰富度似乎低于其他肠型,但疾病并不比其他肠型更严重。
我们还分析了优势菌群与存活脓毒症患者临床指标之间的相关性。分析显示,OTU773(B.vulgatus)和OTU822(Rikenellaceae)与ICU住院时间显著正相关(图4D)。此外,在ICU期间出现并发症的患者OTU773负荷明显较高,但OTU808(E.faecium)负荷没有差异(图4E)。OTU773在脓毒症死亡的患者中更丰富(图4F)。这些结果提示B.vulgatus与脓毒症患者预后不良相关。其他细菌如微球菌科的Akkermansia和OTU40,可能对脓毒症患者有益。OTU40与APACHE II评分呈负相关,而Akkermansia属(已知对许多人类疾病具有保护作用)的丰度在未存活的脓毒症患者的肠道微生物群中较低。
对肠道代谢物与脓毒症疾病严重程度之间相关性的可靠性评估发现,有三种化合物与ICU住院时间呈负相关(图4G)。此外,我们观察到两种与脓毒症相关的代谢物在非存活患者中减少。五种代谢物中有四种是芳香化合物,参与氨基酸代谢,特别是色氨酸和酪氨酸的代谢。5-羟基吲哚乙酰甘氨酸也与APACHE II评分呈正相关,揭示了肠道衍生代谢物与脓毒症之间复杂的相互作用网络(图4G)。我们还观察到,未存活的患者具有丰富的犬尿氨酸(也是色氨酸代谢物)丰度,但在验证组中未检测到。我们观察到这些色氨酸代谢物的丰度在接受不同数量抗生素的患者中保持可比性,表明色氨酸代谢的扰动可能与使用的抗生素数量无关。研究结果总体表明,色氨酸代谢物在脓毒症进展中起着关键作用。
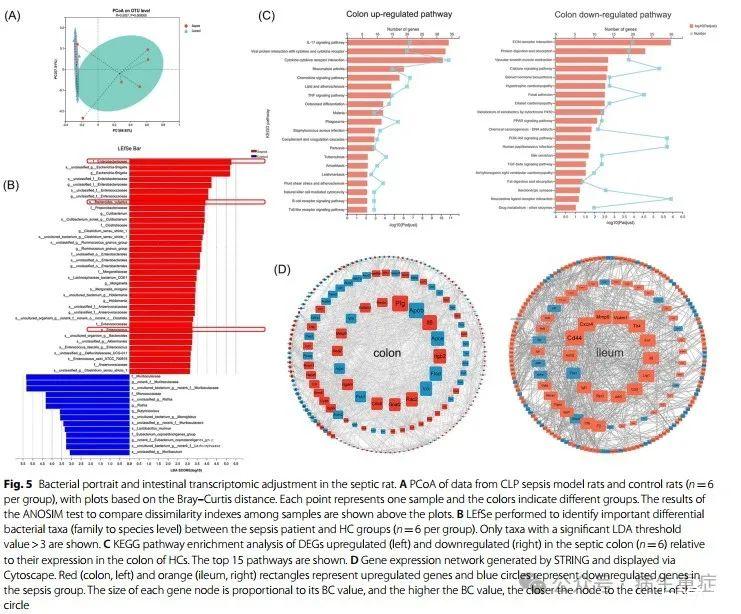
4、脓毒症大鼠肠道菌群和肠道基因表达的变化
即使我们对粪便标本的评估与临床实践中经常遇到的标本类型非常接近,理论上也很难消除临床各种药物对人体肠道微生物群的干扰。因此,我们试图在动物模型中证实脓毒症队列患者的脓毒症相关微生物特征。我们观察到腹腔感染脓毒症患者与肺部感染脓毒症患者具有相似的微生物组结构,我们在大鼠中建立了由腹腔感染引起的经典脓毒症CLP模型。与患者组和HC组相似,脓毒症大鼠组肠道微生物组成与假手术组存在差异(图5A)。同样,在脓毒症大鼠中,必需益生菌(包括Muribaculaceae和Butyricicoccus)的比例下降,而机会致病菌(如Enterobacteraceae和Enterococcus)的比例则增加(图5B)。值得注意的是,尽管脓毒症组患者和HC组的粪便标本中B.vulgatus的负荷相当,但脓毒症大鼠肠道中B.vulgatus的丰度有所增加,B.vulgatus的丰度与脓毒症患者的不良预后有关。
为了进一步阐明脓毒症相关关键细菌的功能,我们评估了细菌丰度谱与肠道基因表达谱之间的关系。最初,我们重点研究了脓毒症大鼠肠道转录组的变化。通过KEGG分析来确定DEGs的功能富集(图5C)。对于回肠和结肠,发现大量在脓毒症中上调的基因参与了细胞因子-细胞因子受体相互作用和趋化因子信号通路,反映在免疫激活上。多种细胞粘附分子在脓毒症结肠中下调,表现为细胞外基质(ECM)受体与转化生长因子-β信号通路相互作用的功能减弱,这有助于免疫细胞的募集和肠道通透性的增强。另一个显著的变化是脂质代谢途径。许多下调的基因与类固醇激素的生物合成、过氧化物酶体增殖物激活受体(PPAR)信号通路和胆固醇代谢有关。胆固醇是几种具有抗炎功能的激素的前体,脂肪酸及其衍生物可激活PPAR信号通路,有效改善脓毒症的炎症损伤。此外,与外源性药物代谢相关的通路在脓毒症中也减弱了。该通路通过肠道代谢物激活芳烃受体(AhR)和孕烷X受体(PXR),随后与核因子-κB信号转导相互作用,发挥抗炎功能。
接下来,通过STRING构建DEGS的蛋白质相互作用表达网络。图5D显示了四个中心DEG。关于结肠中的核心DEG,PLG协助ECM破坏和中性粒细胞迁移,IL6参与炎症,ApoB和APOE是脂蛋白载体的组成部分,可能通过结合内毒素或其他毒素来调节免疫过程。关于回肠中的DEG,CD44、Vcam1和CXCR4服务于免疫细胞聚集过程,而MMP9降解黏附物质以促进细胞运动。综上所述,脓毒症的肠道转录组揭示了与各种类型的宿主反应相关的炎性风暴。
5、脓毒症大鼠肠道转录组中特定细菌的不同相互作用
之前,我们提出拟杆菌和肠球菌是重要的脓毒症相关细菌,可能对疾病进展有不同的影响。因此,我们对微生物丰度和肠道基因表达进行了相关性分析,以确定肠道微生物在脓毒症的病理生理学中的潜在作用。结肠的结果如图6所示。我们能够区分出三个细菌簇(簇1-3)和三个基因簇(簇A-C)。簇1主要包含肠球菌和肠杆菌科,与簇A的基因呈正相关,这些基因在炎症反应(Cybb、Tlr2)和ECM破坏(Mmp13、Spp1)中发挥作用。此外,来自肠杆菌科的簇1生物与与脂质代谢相关基因(Apoc3,Apob)的关系比来自肠球菌的基因更密切。与簇1相比,簇2表现出完全不同的关联趋势。拟杆菌属物种聚集在簇3中,与簇B中的一些基因发生了显著的相互作用。在这些基因相互作用中,Mmp9是一种参与屏障破坏的、研究较为深入的金属蛋白酶,与拟杆菌属物种呈正相关,而编码胱硫氨酸-γ-裂解酶的Cth,则与拟杆菌属物种呈负相关,Cth是一种主要的内源性硫化氢产生酶,通过IL-8信传导抑制炎症,呈负相关。

讨论
研究亮点
使用微生物组和非靶向代谢组学的组合来分析入院时脓毒症患者的粪便样本,然后筛选出可能在疾病结局中发挥重要作用的微生物群,代谢物和潜在的信号通路。最后,通过脓毒症动物模型的微生物组学和转录组学分析验证了上述结果。
局限性
首先,研究的样本量较小,所有参与者都来自同一个医疗中心;因此,我们的结果应该在其他机构的患者中得到验证。其次,尽管我们的研究结果在控制年龄时仍然显著,并在大鼠模型中得到验证,但临床混杂因素仍然存在,包括入ICU前的药物治疗和感染的病原体。第三,肠球菌和普通双歧杆菌对脓毒症的影响是通过相关性分析推断出来的,应该通过对特定细菌定植的动物实验来验证。未来的研究应使用宏基因组学技术来精确揭示与脓毒症肠道代谢物改变有关的微生物遗传元素的功能。
相关新闻推荐
1、不同pH、Na2CO3、NaCl浓度对耐盐碱菌株NM-1生长曲线的影响——材料与方法
2、微生物生长曲线绘制,Bioscreen可以取代这两个传统组合仪器
3、芽孢杆菌对黄曲霉毒素B1、呕吐毒素、玉米赤霉烯酮的降解作用——材料与方法