通过将一个物种添加到“生态背景”中的“功能效应”与背景群落的功能联系起来,可以很好地反映该物种的“功能效应”。为了进一步拓展研究范围,探索微生物群落功能的全局上位效应这一现象是否具有普遍性,作者重新分析了多个已发表的数据集,检验全局上位效应在不同生态环境中的适用性。作者通过分析细菌、植物和浮游植物的群落,确认这些不同生态系统中的群落功能也可以通过全局上位效应的模型进行预测(图2)。每个数据集涉及不同的物种组合、生态条件和群落功能(如生物质生产、代谢活动、酶分泌等),但简单的统计模型能够解释各个数据集中约75%的功能效应方差。可以看到,大多数物种(约50%,红线)的FEE呈负斜率,即我们在背景介绍中提及的“收益递减”或“成本增加”现象,这种在群体遗传学中也十分常见。而少数物种(约5%,蓝线)的FEE呈正斜率,即在功能较高的背景中加入该物种会更有益(或更无害)。
其余的(约45%,黑线)则表现出趋近于零的斜率,尽管如此,其FEE的残差仍有助于我们辨别:(1)残差较小的物种:即其对群落功能的贡献很大程度上独立于其生态背景;(2)残差较大的物种:即其对群落功能的贡献较大程度上取决于群落背景的物种组成(图2)。这些结果表明,全局上位效应具有广泛的适用性,不仅在微生物群落中可行,还可以应用于更大规模的生态系统中。
通过这些不同生态环境的研究,作者确认了全局上位效应在生态学中的普遍性,这为未来群落功能预测提供了更广阔的视角。

图2焦点物种的功能效应通常可以通过将其与其所属背景群落的功能联系起来的简单回归来预测(红线:负斜率,蓝线:正斜率,黑线:接近零的斜率),在由非常不同的生物类型、不同的生态条件和不同的集体功能组成的群落中,都观察到了这一点全局上位效应已被证实在不同的生态系统中存在,但其具体的形成机制仍不明确。作者通过分析物种间的成对相互作用,试图揭示这些相互作用如何累积并影响群落的整体功能表现,从而更好地理解物种相互作用的微观机制与群落功能之间的联系。
作者首先提出了一个核心假设:全局上位效应模型中回归斜率可以用物种之间的成对相互作用量化。具体而言,他们认为每个物种对群落功能的贡献,不仅取决于该物种本身的特性,还和它与群落中其他物种之间的相互作用密切相关。因此,群落功能效应方程(FEE)中的回归斜率可以通过这些物种间相互作用的总和来解释。为了验证这一假设,作者设计了一套分析流程。首先,他们定义了“有效相互作用”(effective interactions),这是指两个物种之间的功能相互作用被平均之后的影响:
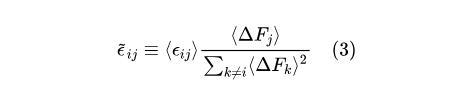
其中<εij>表示物种i和j之间的平均功能相互作用,即两种物种共同存在时,与各自独立功能效应的加性期望相比,其在群落中的平均功能差异(图3B),而<ΔFj>则表示物种j在不包括物种i的生态背景下的平均功能效应。基于数量遗传学中的研究结果,可以推断:FEE中物种i加入后(对应数量遗传学中的突变i)的斜率bi可以近似为其他所有物种j(突变j)的“有效相互作用”的和(图3C)。简单来说,有效相互作用量化了每对物种在不同生态背景下如何共同影响群落的整体功能。通过这种方式,他们能够量化不同物种在群落中的功能贡献,并探索这些贡献是如何随着物种间的相互作用而变化的。
为了验证这一理论,作者在前文提及的实验与数据集中测试并计算了物种间的有效相互作用,即通过分析每对物种在所有可能的背景下的功能变化,得出它们之间相互作用的平均效应。通过对实验数据的深入分析,作者发现,物种间的成对相互作用确实能够解释功能效应方程中回归斜率的变化(图3D)。这意味着,一个物种对群落整体功能的贡献不仅仅是孤立的,而是受到其与其他物种相互作用的影响(请注意,这里的相互作用是指物种对在所有生态背景下对群落水平功能的平均影响,并不一定对应于两个物种之间的直接正向或负向生态相互作用)。例如,他们发现,具有强烈负相互作用的物种对群落功能的贡献往往在具有高背景功能时显著下降,而具有正相互作用的物种则在背景功能较高时能够进一步提升群落功能(图3E-G)。
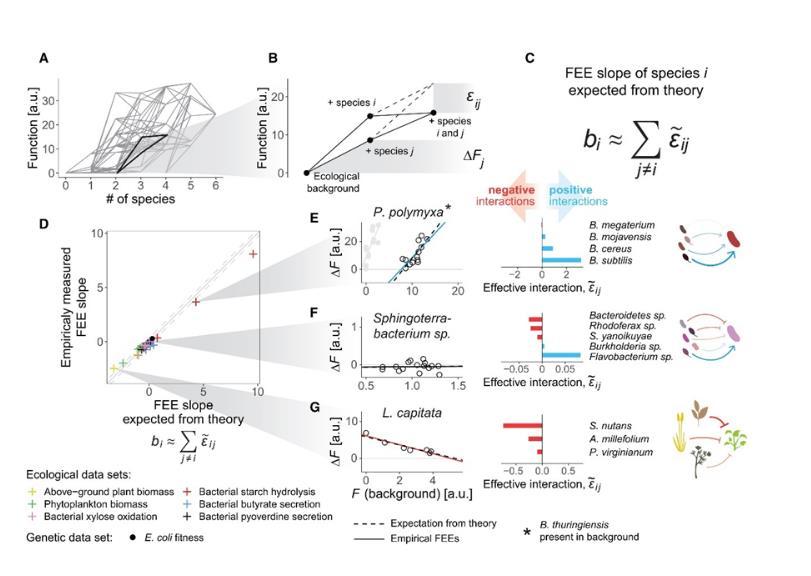
图3(A)Sanchez-Gorostiaga等人实验中物种数量与群落功能之间的映射。每个节点代表一个联合体,边用于连接由不同物种组成的联合体;(B)包含两个物种的示例(i和j)在特定生态背景下导致功能(实线)低于加性期望(虚线)。这种差异(εij)表示两个物种之间的功能有效相互作用(直接相互作用或间接相互作用);(C)斜率基于数量遗传学的理论结果;(D)量化每个物种和数据集的所有有效相互作用,估计预期的FEE斜率,估计的斜率与图2中的经验拟合结果一致;(E-G)三种斜率分别为正斜率、零斜率和负斜率的物种,斜率由焦点物种与其生态伙伴之间的有效相互作用的符号和大小来描述。
有趣的是,作者还利用这一理论为数据集研究中发现的反常现象进行了解释:在图2D中作者观察到了多黏芽孢杆菌(P.polymyxa)的FEE在苏云金芽孢杆菌(B.thuringiensis)存在与否的背景下表现出了两个分支,通过有效相互作用方程可以得知,多黏芽孢杆菌与苏云金芽孢杆菌表现出了强烈的负相互作用,而与其他物种则都表现出了正相互作用。生态背景中苏云金芽孢杆菌的存在与否决定了在FEE中观察到的两个分支,而这种现象也可以从分子机制上得到解释:多黏芽孢杆菌是一种生物素(Biotin)营养缺陷型物种,其生长由群落中其他成员的交叉喂养(Cross-feeding)来促进,这一定程度上说明了定义物种间有效相互作用的实用性。
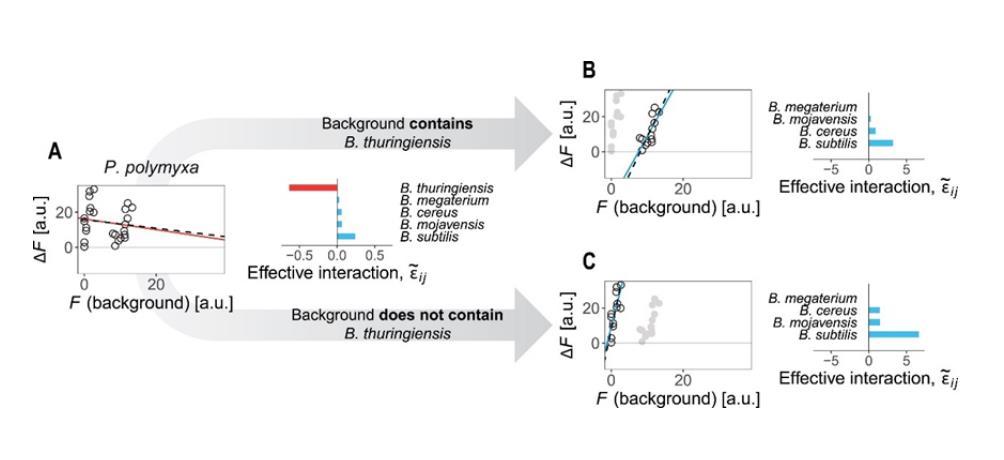
图S14(A)当为所有物种背景拟合单个FEE时,可以观察到负斜率,这可以通过多黏芽孢杆菌与苏云金芽孢杆菌之间的强烈负有效相互作用来解释;(B-C)当苏云金芽孢杆菌存在或不存在时,多黏芽孢杆菌与其余物种的有效相互作用为正值,这导致每个分支中FEE的斜率为正。虚线表示估算的FEE,实线是数据的经验性拟合结果
通过计算和验证物种间的有效相互作用,作者成功揭示了全局上位效应的形成机制。他们的分析表明,群落功能是由物种间复杂的相互作用网络共同决定的,而这些相互作用可以通过简单的统计模型加以预测。这一发现进一步证明了全局上位效应不仅适用于基因层面的相互作用,也同样适用于生态系统中的物种间相互作用。
为了进一步探讨如何利用物种间的相互作用进行实际的群落功能预测,作者利用功能效应方程(FEEs)构建一个可操作的框架,以准确预测群落中不同物种组合对整体功能的影响。
作者提出,FEE可以用于预测特定物种组合对群落整体功能的影响。具体实现的思路是,通过逐步累加每个物种的功能效应,来推断一个新的群落组合的功能表现。通过先预测一个已知群落的功能表现,然后逐步加入新的物种,利用FEE累加预测其功能的增减效应,从而得出目标群落的最终功能:
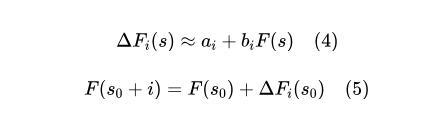
由于群落功能的预测涉及多个物种的组合,作者提出了一个迭代方法,即将每个物种的FEE与之前的群落功能预测结果进行相加(图4A)。由于方程(2)中残差项θi(s)的存在,物种加入的顺序可能会影响最终的预测结果,作者通过一种统计方法解决了这一问题,使得物种不同组装顺序下的功能预测结果一致。
为了检验这一预测框架的准确性,作者在实验中对61个之前没有组装过的土壤细菌群落进行了预测,并将预测结果与实验观测结果进行比较。结果表明,模型能够准确预测这些群落的功能,相关系数达到0.80(图4B),展示了其在实际应用中的高效性。这个结果表明,FEE不仅能够描述已有数据中的功能效应,还能成功预测未观测到的新组合体的功能。
除此之外,作者在预测群落最佳功能组成的测试中也得到了相较于其他统计模型更精确的结果以及具有最佳功能的群落组成。这对于微生物共生体的设计和优化,特别是在生物技术应用中,如生物燃料生产或污染物降解等,具有重要的现实意义。
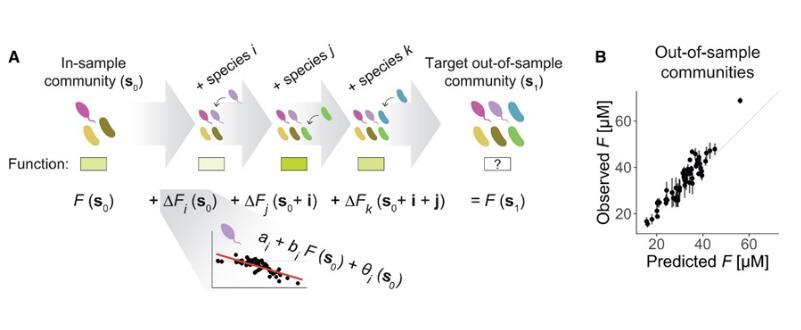
图4(A)假设连接一组物种(这里指物种i,j和k)的功能效应,从而基于样本内群落预测样本外群落;(B)通过构建61个新的合成群落并比较预测和观测的Pyoverdine分泌水平来评估该方法的性能,可以发现观测值域预测值之间具有良好的一致性
总而言之,本文讨论了如何通过数量遗传学中全局上位效应的生态学类比来克服微生物群落功能预测中的挑战。目前,工程化微生物群落在生产生物燃料、降解污染物和拮抗病原体等领域的应用越来越受到关注。然而,构建能够整合复杂功能和生态交互的群落功能预测模型仍然十分困难。通过功能效应方程(FEEs)可以建立简单、可扩展的线性模型,预测物种在特定背景群落中的功能效应。这种模型不仅适用于多种生态系统,还不需要逐一解析每对物种之间的复杂分子机制。研究表明,FEEs能有效简化群落功能预测,并提供一种新的方式来优化群落功能。这一方法为开发更先进的预测工具提供了基础,推动了微生物群落工程在实际应用中的前景。
文章亮点
1.提出了全局上位效应可扩展到群落生态学的假设,并通过实验验证了其普遍性;2.创新地通过有效相互作用量化了物种对群落功能的贡献,揭示了功能效应的机制;3.开发了一种基于功能效应方程(FEE)的预测工具,能够精确预测未来物种组合的功能表现;4.FEE模型不仅能够预测群落功能,还能识别出功能最优的物种组合,为实际应用提供了指导。
相关新闻推荐
3、生长曲线和杀菌曲线试验验证去甲泽拉木醛联合美罗培南杀菌效果