2.4山梨醇高渗胁迫对玉米大斑病菌菌丝形态的影响
山梨醇高渗胁迫对玉米大斑病菌菌丝形态的影响结果见图3。由图3可以看出,在无山梨醇添加的DPA培养基上,WT菌株和KO菌株的菌丝形态无明显差别。在1.0 mol/L山梨醇胁迫条件下,KO菌株的菌丝形态没有明显变化,而WT菌株的菌丝细胞内出现了大量的小型颗粒状物质。在1.5 mol/L山梨醇胁迫条件下,KO菌株的菌丝细胞内也出现了大量的小型颗粒状物质,而WT菌株的菌丝细胞内出现了较多的大型颗粒状物质。在2.0 mol/L山梨醇胁迫条件下,KO菌株的菌丝变粗,原生质体浓缩成颗粒状,而WT菌株的菌丝细胞内出现很多空隙。
2.5山梨醇高渗胁迫对玉米大斑病菌菌丝细胞内脂类物质沉积的影响
由于脂类物质是玉米大斑病菌菌丝内主要的渗透胁迫调节物质[13],在图3中出现的颗粒状物质极有可能是脂类物质的积累。因此,本研究利用油红O染色剂,对WT菌株和KO菌株在2.0 mol/L山梨醇高渗胁迫条件下菌丝细胞内脂类物质的沉积情况进行显微观察,结果(图4)发现,与高渗胁迫处理前相比,2.0 mol/L山梨醇高渗胁迫后,WT菌株菌丝细胞内的脂滴数量明显增多,并且脂滴体积明显增加,而KO菌株的菌丝细胞变粗膨大,脂滴数量有所减少,但脂滴体积显著变大。脂滴的分布情况也明显不同,在高渗胁迫处理前,WT菌株和KO菌株的脂滴分布都不均匀,但山梨醇高渗胁迫处理后,WT菌株的脂滴大小及分布都比较均匀一致,而KO菌株大脂滴和小脂滴并存且分布极不均匀,位于中部或偏于某一侧。该试验结果与图3的表现基本一致。
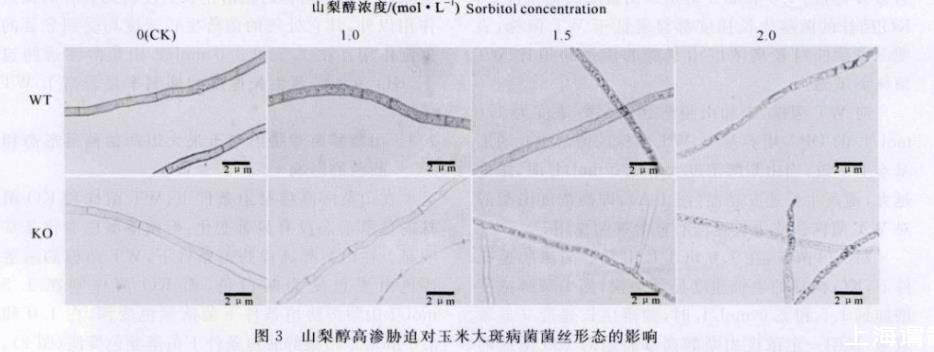
图3山梨醇高渗胁迫对玉米大斑病菌菌丝形态的影响
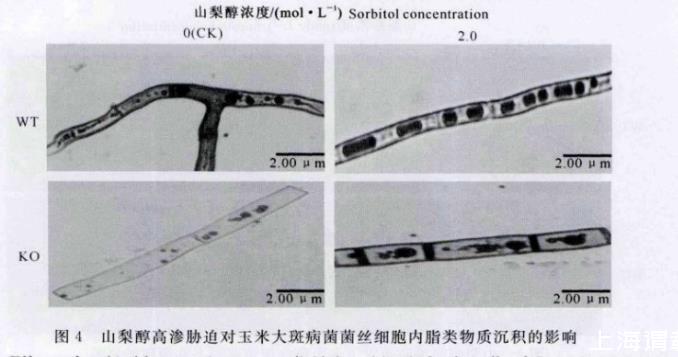
图4山梨醇高渗胁迫对玉米大斑病菌菌丝细胞内脂类物质沉积的影响
3讨论与结论
山梨醇是一种重要的渗透胁迫调节物质,它可以与水、甘油和丙二醇混溶,因此具有与NaCl、KCl等盐分子不同的化学性质[18]。在植物的耐盐胁迫研究中,200μg/mL山梨醇浸种处理可促进无花果种子萌发,加速可溶性糖和脯氨酸积累,增强无花果幼苗渗透调节,有效降低盐胁迫对幼苗细胞造成的伤害,提高无花果幼苗耐盐性[19]。因此,山梨醇渗透胁迫与盐胁迫具有不同的渗透调节机制。
在本课题组的前期研究中发现,在相同的盐离子浓度条件下,与野生型(WT)菌株相比,STK1基因敲除突变体(KO)菌株的菌丝生长缓慢,细胞内容物减少,生理状况降低[12,14]。但是本试验的研究结果表明,在1.0 mol/L山梨醇渗透胁迫条件下,玉米大斑病菌KO菌株生长状况与WT菌株没有显著差异,只有当山梨醇浓度提高到1.5 mol/L以上时,才显著抑制了KO菌株的菌丝生长速度。
因此,STK1基因在山梨醇高渗胁迫调节过程中存在一个滞后的信号应答过程。另外,与非渗透胁迫相比,在1.0 mol/L山梨醇渗透胁迫条件下,KO菌株菌丝生长速度不但未受到抑制,反而得到促进,说明1.0 mol/L山梨醇处理使玉米大斑病菌菌丝恢复了部分渗透胁迫调节功能,即1.0 mol/L山梨醇处理可能增强了菌丝的渗透胁迫调节能力,“补偿”了STK1被敲除后造成的某些功能障碍。该试验结果与山梨醇是桃[20]、烟草[21]等植物渗透调节物质和医药领域渗透剂[22]的结论是一致的,并且Stavniichuk等[23]研究表明,山梨醇通路与MAPK通路可协同调控糖尿病的发生。
但是,更高浓度的山梨醇对这一未知途径也产生了一定的阻遏作用,使玉米大斑病菌的渗透胁迫调节能力受到了不可逆的抑制作用,不仅菌落生长速度受到抑制,而且细胞壁色素的合成受到干扰,菌丝颜色变浅;细胞内物质的降解能力下降,出现颗粒状物质积累。研究表明,参与渗透胁迫调节的细胞壁色素物质为黑色素,而这些颗粒状物质极有可能是渗透胁迫调节物质[24-25]。王梅娟等[13]研究表明,玉米大斑病菌的渗透胁迫调节物质主要为甘油,而真菌细胞内甘油主要来自于脂肪的降解。因此,菌丝细胞内脂类物质沉积形成的脂滴大小和分布与渗透胁迫调节能力密切相关。在本试验中发现,山梨醇高渗胁迫条件下,脂类物质的沉积与颗粒状物质的分布基本一致。利用油红O染色观察脂类物质在菌丝内的沉积情况,发现STK1基因缺失后脂滴的大小及分布均与野生型菌株不同,因此,STK1基因很可能通过调控脂类物质合成进行渗透胁迫调节。