2.4.4致病菌生长与碳源的关系培养3 d后,菌落直径与不同碳源的关系如图8所示。
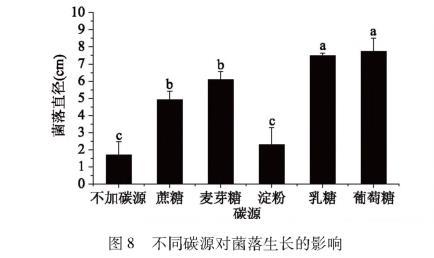
图8不同碳源对菌落生长的影响
由图8可以看出,在不加碳源以及以蔗糖、麦芽糖、淀粉、乳糖、葡萄糖为碳源时,该菌均能生长,但是各实验组菌落直径存在显著差异。不加碳源的培养基菌落直径增长最慢;以淀粉为碳源时,菌落直径与不加碳源组差异不显著、与其他组有显著性差异;以蔗糖和麦芽糖为碳源时,两组菌落直径差异不显著,菌落直径相对较大,但与葡糖糖和乳糖为碳源时的菌落直径有显著差异,且比这两组的菌落直径要小;以葡萄糖和乳糖为碳源时,两组菌落直径差异不显著,但葡萄糖的菌落直径明显要比乳糖大。综上,碳源对该致病菌生长有重要影响,在以葡萄糖为碳源时该菌长势最好,而以蔗糖、乳糖、麦芽糖这些双糖作为碳源时生长稍差,以淀粉为碳源时生长最差。
2.4.5菌丝生长与氮源的关系
相同的培养时间,菌落直径与不同氮源的关系如图9所示。
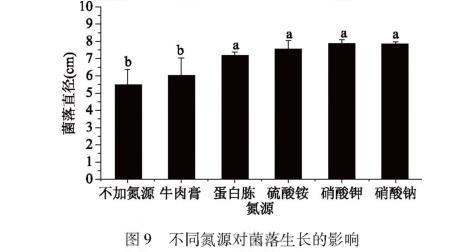
图9不同氮源对菌落生长的影响
由图9可以看出,不加氮源实验组以及以牛肉膏、蛋白胨、硫酸铵、硝酸钠、硝酸钾为氮源的实验组菌落均能生长,但是不加氮源组的菌落直径明显落后于加氮源组的,说明氮源为该菌生长的重要条件。从图中可以看出,蛋白胨组、硫酸铵组、硝酸钾组和硝酸钠的菌丝直径差异不显著,但硫酸铵的菌落直径>蛋白胨菌落菌落直径,而以硫酸铵为氮源的菌落直径明显低于以硝酸钠、硝酸钾为氮源的实验组。综上,氮源为该致病菌生长的重要影响因素,且以硝酸钠、硝酸钾为氮源时,该菌生长最快,因此硝酸钠、硝酸钾类硝酸盐类为该致病菌生长的最佳氮源。
2.5致病菌致死温度区间的确定
分别经35、40、45、50、55、60℃水浴20 min并振荡培养72 h后,菌丝生长情况如图10,回接后菌丝生长情况如图11。
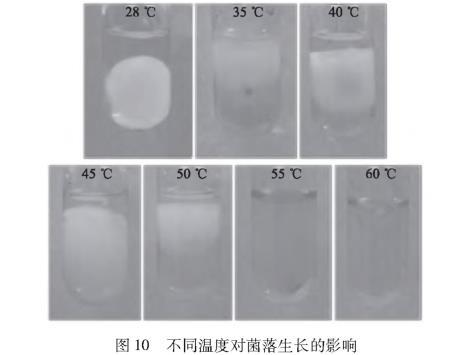
图10不同温度对菌落生长的影响
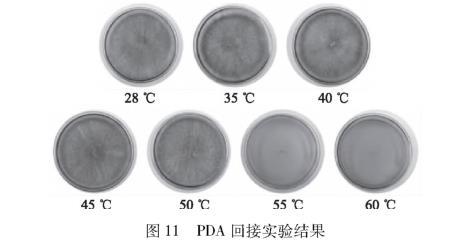
图11 PDA回接实验结果
由图10可知,分别经35、40、45、50、55、60℃水浴20 min后,28℃对照组、35、40、45、50℃实验组均有菌丝生长且均形成了菌丝球,55、60℃的PDB培养液清澈,无菌丝生长。
取各试管中的培养液进行PDA回接实验,结果如图11。经28、35、40、45、50℃水浴20 min后回接在PDA培养基上,PDA培养基均有菌落生长,而经55、60℃水浴20 min后回接在PDA培养基上,PDA培养基无菌落生长。由以上实验可知:该致病菌的致死温度区间为50~55℃。
3讨论与结论
本实验以山西榆次樱桃园流胶树体上分离纯化得到的一株葡萄座腔菌为试验菌株,对其生物学特性进行了试验研究。预实验阶段将葡萄座腔菌分别接种于查氏培养基和PDA培养基,结果发现PDA培养基更适合此菌生长;试验阶段通过对菌丝结构的观察,发现该致病菌菌丝有明显节状结构,内含横膈膜,同时菌丝有树状分支,有些菌丝联结成网状。通过菌落划伤、近紫外线照射及营养缺乏进行诱导产孢试验发现,三种胁迫方式均能诱导其产生孢子,这一观察与何靖柳等在观察红阳猕猴桃葡萄座腔菌时的结果相反,却与韩青梅等在研究Botryosphaeriadothidea侵染苹果果实的研究结果一致。
该葡萄座腔菌的孢子为卵形或椭圆形,大小不一,有聚集现象,与Phillips等对葡萄座腔菌孢子形态的描述一致。
研究最适生长条件的实验结果表明,温度、光照、pH、碳源、氮源均为葡萄座腔菌生长的重要影响条件。葡萄座腔菌最适温度为28℃,有力说明了5~9月份为发病较为严重阶段的原因。葡萄座腔菌最适光照条件为12 h光照/12 h黑暗交替。卢伟对桂花叶斑病病原葡萄座腔菌(B.dothidea)进行生物学特性的研究试验也认为病原菌生长的最适光照条件为交替光照,但罗光明等认为光照对病原菌的生长无明显影响。葡萄座腔菌最适生长pH为6.0,说明致病菌适生存于偏酸性环境。最适碳源为葡萄糖,最适氮源为硝酸钠、硝酸钾类硝酸盐。该致病菌水浴处理20 min的致死温度区间为50~55℃,这与黄静等对侵染草莓的落叶松葡萄座腔菌的致死温度70℃(20 min)不一致;这说明不同植株上分离的致病菌虽然同属葡萄座腔菌,但生物学特性有所差异。这些结果表明即使是同类植物,分离得到的致病菌特性也不相同,因此对葡萄座腔菌应该专株专治。
我国多样的气候及各种地理环境造就了丰富的葡萄座腔菌科真菌。本实验所用的葡萄座腔菌来自山西榆次樱桃园流胶树体,葡萄座腔菌是引起多种树体流胶的病原菌之一,通过本实验对该致病菌生物学特性的研究结论,可以为其防治工作提供一定的理论基础:调节樱桃树的栽种温度,栽种土壤中性偏碱,避免硝酸类肥料,在樱桃树生长过程中要保护树体,避免让树体出现伤口,减少紫外线照射,尽可能破坏致病菌生长的最佳条件,抑制葡萄座腔菌的生长,从而降低甜樱桃流胶病的发病率,增加甜樱桃产量。